
POTENTIAL ZOONOTIC BAT-BORNE DISEASE IN INDONESIA (Part 2)
As an archipelago, Indonesia’s seas and oceans, in theory, may have historically. Globalization and resulting changes in human activities, including wildlife trade, are increasing across and beyond the country, heightening the risk of cross-species transmission and the spread of pathogens. In addition, the potential for diseases to “spillback” into animals from humans also enables greater potential pathogen spread and poses concerns for biodiversity conservation (1–5).
Given Indonesia’s abundant biodiversity of natural reservoir hosts for viruses (e.g., non-human primates, rodents, and bats), high tropical deforestation rates, wildlife trade and hunting networks, and growing human population, the risk for zoonotic disease emergence is high. These rapid ecological changes are bringing humans into close contact with wildlife species that were previously rarely seen (6). It is probable that those factors may increase the risk for zoonotic disease emergence, and therefore the country should become a hotspot in targeting surveillance to identify spillover events.
In the last edition, we have explained the potential presence of coronavirus circulating in Indonesia, which may have spilled over into human populations several times within the region but are either not reported or otherwise missed by clinical surveillance (17). This edition will focus on other virus families that can also be a threat to become zoonotic outbreaks in Indonesia.
Filovirus
To date, 12 distinct filoviruses have been described. The seven filoviruses that have been found in humans belong either to the genus Ebolavirus (Bundibugyo virus (BDBV), Ebola virus (EBOV), Reston virus (RESTV), Sudan virus (SUDV) and Taï Forest virus (TAFV) or to the genus Marburgvirus (Marburg virus (MARV) and Ravn virus (RAVV)). Among the filoviruses family, the filoviruses Ebola virus (EBOV) and Marburg.
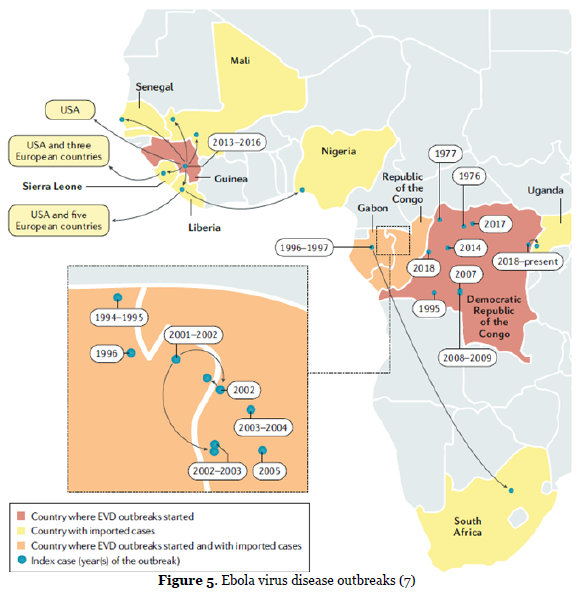
virus (MARV) are defined as category A pathogens by the NIH (https://www.niaid.nih.gov/research/emerging-infectious-diseases-pathogens) and the Centers for Disease Control and Prevention (CDC; https://emergency.cdc.gov/agent/agentlist-category.asp). There were two major EBOV outbreaks in the last decade. The 2013–2016 epidemic was primarily in West African countries, infecting ~30,000 individuals with a mortality rate of 40%. There was a second large outbreak in the Democratic Republic of the Congo from 2018 to 2020, with around 3,500 infections and a mortality rate of 65%. Until 2020, ~33,604 EBOV infections in humans, including 14,742 deaths (average CFR 43.8%) are on record. Bats are the primary reservoir for EBOV, and the virus can transmit either directly to humans or through intermediate zoonotic hosts. It is thought that fruit bats of the Pteropodidae family are natural EBOV hosts. Once in the human population, EBOV can spread through blood and bodily fluids or sexual transmission (7).

It can be difficult to clinically distinguish EVD from other infectious diseases such as malaria, typhoid fever and meningitis. Ebola is vastly less contagious (compared to SARS), evidence to date suggests that transmission of Ebola occurs only through direct contact with bodily fluids, and furthermore, only after an individual has become symptomatic. EVD has a high case–fatality rate (about 50%); it is characterized by fever, gastrointestinal signs and multiple organ dysfunction syndrome (7).
Despite the discovery of EBOV (Reston virus) in nonhuman primates and domestic pigs in the Philippines and the serological evidence for its infection of humans and fruit bats, information on the reservoirs and potential amplifying hosts for filoviruses in Asia is lacking. In a study, serum samples collected from 353 healthy Bornean orangutans (Pongo pygmaeus) in Kalimantan Island, Indonesia, during the period from December 2005 to December 2006 were screened for filovirus-specific IgG antibodies using a highly sensitive enzyme-linked immunosorbent assay (ELISA) with recombinant viral surface glycoprotein (GP) antigens derived from multiple species of filoviruses (5 EBOV and 1 MARV species). They showed that 18.4% (65/353) and 1.7% (6/353) of the samples were seropositive for EBOV and MARV, respectively, with little cross-reactivity among EBOV and MARV antigens. In these positive samples, IgG antibodies to viral internal proteins were also detected by immunoblotting. Interestingly, while the specificity for Reston virus, which has been recognized as an Asian filovirus, was the highest in only 1.4% (5/353) of the serum samples, the majority of EBOV-positive sera showed specificity to Zaire, Sudan, Cote d’Ivoire, or Bundibugyo viruses, all of which have been found so far only in Africa. These results suggest the existence of multiple species of filoviruses or unknown filovirus-related viruses in Indonesia, some of which are serologically similar to African EBOVs, and transmission of the viruses from yet unidentified reservoir hosts into the orangutan populations (8). These findings point to the need for risk assessment and continued surveillance of filovirus infection of human and nonhuman primates, as well as wild and domestic animals, in Asia.
Henipavirus
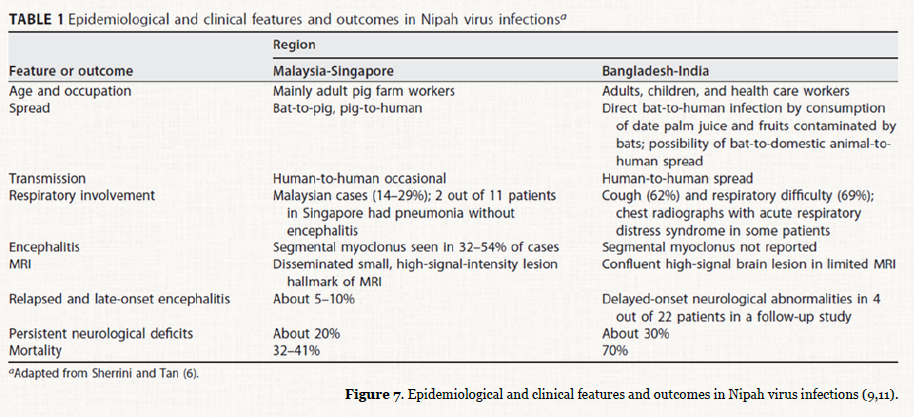
In henipaviruses, a genus in the family Paramyxoviridae, >350 human fatalities from Hendra (HeV) or Nipah virus (NiV) disease outbreaks have been reported. Nipah Virus (NiV) was discov-ered in 1998 during the first reported outbreak in the Sungai Nipah village in Malaysia. During that initial outbreak, 283 hu-man cases of acute encephalitis were diagnosed (mainly in farmers), with 109 deaths. Clinical features of NiV disease include acute respiratory distress and severe encephalitic symptoms, including seizures, convulsions, and coma. The onset of disease is very abrupt, and the course of the severe phase is very brief. Since that first outbreak, no other outbreaks of NiV disease have been detected in Malaysia. However, 2 years later, NiV was detected in both Bangladesh and India. Since 2001, sporadic outbreaks have occurred in Bangladesh almost every year, mainly detected retrospectively by the established countrywide encephalitis surveillance program. NiV encephalitis is still a rare disease, involving <5% of the total number of reported encephalitis cases investigated in Bangladesh. NiV is a single-stranded, negative-sense RNA virus of approximately 18.2 kb long, belonging to the family Paramyxoviridae. NiV can be transmitted to humans from animals (such as bats or pigs), or contaminated foods and can also be transmitted directly from human-to-human. The animal host reservoir for NiV is the fruit bat (genus Pteropus), also known as the flying fox (9–11).
Sequence analyses of the viral genomes from the Malaysian and Bangladesh/India outbreaks showed that NiV separates into 2 distinct lineages/genotypes: Nipah-Malaysia (NiV-M) and Nipah-Bangladesh (NiV-B). In addition to differences in genetics and geographic distribution, the viral lineages differ in several other important ways. First, while infected pigs acted as the intermediate host for NiV-M, no intermediate host has been identified for NiV-B. In contrast to NiV-M, the primary source of NiV-B human infections is the consumption of raw date palm sap contaminated by virus shed by infected fruit bats. Human-to-human transmission was not clearly observed during the NiV-M outbreak but was well documented in a number of NiV-B outbreaks. In addition, case-fatality rates for NiV-B outbreaks have been much higher (range, 60%–100%) than those caused by NiV-M (39%). However, differences in mortality rates may be attributable to differences in healthcare support in the different countries and the fact that Bangladeshi outbreaks are usually identified retrospectively. Finally, in addition to the encephalitic symptoms seen during NiV-M infection, acute respiratory distress symptoms are also seen in cases of NiV-B infection (9–11).
Compared to other viruses, research on the Nipah virus has been limited in Indonesia because attributable disease outbreaks have not been reported. One study reported the detection of Nipah virus genome in in P. vampyrus in Sumatera, Indonesia, between 25 and 29 May, 2009, using real-time PCR. A total of 215 samples (71 oro-pharangeal swabs, 71 blood samples, 32 pooled urine samples and 41 urinary bladder samples) were collected from 71 P. vampyrus flying-foxes from two locations (Kota Medan and Deli Serdang Kampung) in the Indonesian province of North Sumatera. Four samples yielded Nipah virus genome – an oro-pharangeal swab and a bladder sample from DS 21 (an adult female), and two pooled urine samples containing urine from DS 21. Virus isolation was not undertaken. Nipah virus is categorized as a BSL 4 agent, and Indonesia does not currently have a laboratory with BSL4 facilities. Realtime PCR and RT- PCR represent a practical and robust alternative to detect Nipah virus from field samples in this situ-ation. Their analyses showed that the Indonesian and Malaysian nucleotide sequences were more closely aligned that sequences with each other than they were with the Bangladesh or Indi-an sequences (12). This is not unexpected given the demonstrated movement of flying foxes between peninsular Malaysia and Sumatera across a sea distance of less than 50 km. Other study identified unique paramyxovirus sequence from three species of fruit bats (Pteropus vampyrus, Pteropus hypome-lanus and Acerodon celebensis). Fruit bats were captured in: Panjalu District, West Java Province during February, 2010 (n = 26); Lima Puluh Kota District, West Sumatra Province during February, 2011 (n = 20). Their findings are potentially repre-senting three new henipaviruses and two new rubulaviruses among fruit bat populations in Indonesia (13).
Other studies have demonstrated anti-Nipah virus antibodies in flying foxes in Indonesia. One study nonrandomly sampled 106 P. vampyrus bats from market sellers on the Indonesian islands of Java and Sumatra during a 12-day period from July 23 to August 3, 2002. Serum samples from 32 bats neutralized NiV (median titer 20, range 5–160), samples from 52 bats did not, and samples from 20 bats caused toxic reactions in the cell sheet at dilutions <10 (n = 7), <20 (n = 9), or <40 (n = 4), precluding a definitive test outcome. The detection of antibodies that neutralized NiV at all 3 sampling locations indicates that infection with NiV (or a cross-neutralizing virus other than HeV) is widespread in P. vampyrus in Sumatra and Java. These find-ings, in conjunction with earlier findings in peninsular Malaysia, suggest that NiV infection is likely to be found in P. vampyrus across its entire range (14). Other group conducted study of farmer interviews and a serologic survey of 610 pig sera and 99 bat sera from West Kalimantan province. Farmers reported no recent or historic encephalitic or respiratory disease in themselves, their families, workers or pigs. The survey found no evidence of exposure to Nipah virus in pigs. However, serological evidence Nipah virus was detected in 19% of the 84 Large Fly-ing-foxes (Pteropus vampyrus) from West Kalimantan, Borneo. Another study provides evidence for the presence of NiV east of Wallace’s Line in East Timor (Sulawesi, Sumba or New Guin-ea) (15). This study, in combination with the serological evidence of henipavirus infection in P. vampyrus from Sumatra, Java and Borneo has shown that henipaviruses occur in fruit bats widely across the Sunda Shelf, Wallacea and New Guinea (16).
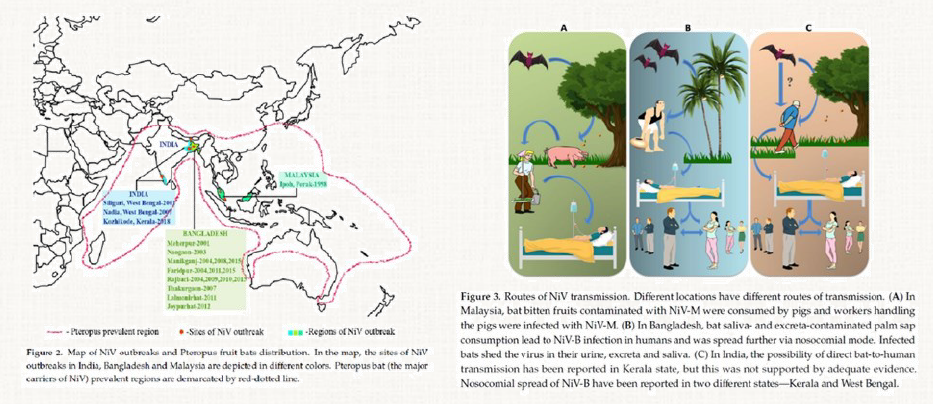
NiV emerged as a new virus exactly 20 years ago, causing severe morbidity and mortality in both humans and animals and destroyed the pig-farming industry in Malaysia, and it contin-ues to cause outbreaks in Bangladesh and India (11). However, serology survey data on evidence NiV exposure to Indonesian people were scarce, although the moderate endemic of NiV in P. vampyrus in Indonesia had been shown by several above studies. As the reservoir host Pteropus bat is widespread, and NiV has been found in bats in various countries, the potential for outbreaks to occur in new regions remains significant (17). Ongoing surveillance is required to detect indicative changes in infection dynamics in fruit bats or the early introduction of infection to the pig and human population.
References
- Letko M, Seifert SN, Olival KJ, Plowright RK, Munster VJ. Bat-borne virus diversity, spillover and emergence. Nat Rev Microbiol [Internet]. 2020;18(8):461–71. Available from: https://doi.org/10.1038/s41579-020-0394-z
- Ruiz-Aravena M, McKee C, Gamble A, Lunn T, Morris A, Snedden CE, et al. Ecology, evolution and spillover of coronaviruses from bats. Nat Rev Micro-biol [Internet]. 2021; Available from: https://doi.org/10.1038/s41579-021-00652-2
- Dharmayanti NLPI, Nurjanah D, Nuradji H, Maryanto I, Exploitasia I, Indriani R. Molecular detection of bat coronaviruses in three bat species in Indonesia. J Vet Sci [Internet]. 2021 Nov;22(6):0. Available from: https://doi.org/10.4142/jvs.2021.22.e70
- Afelt A, Frutos R, Devaux C. Bats, Coronaviruses, and Deforestation: Toward the Emergence of Novel Infectious Diseases? [Internet]. Vol. 9, Frontiers in Microbiology . 2018. p. 702. Available from: https://www.frontiersin.org/article/10.3389/fmicb.2018.00702
- Sánchez CA, Li H, Phelps KL, Zambrana-Torrelio C, Wang L-F, Olival KJ, et al. A strategy to assess spillover risk of bat SARS-related coronaviruses in Southeast Asia. medRxiv : the preprint server for health sciences. 2021.
- Tollefson J. Why deforestation and extinctions make pandemics more likely. Nature. 2020;584(7820):175–6.
- Jacob ST, Crozier I, Fischer WA 2nd, Hewlett A, Kraft CS, Vega M-A de La, et al. Ebola virus disease. Nat Rev Dis Prim. 2020 Feb;6(1):13.
- Nidom CA, Nakayama E, Nidom R V, Alamudi MY, Daulay S, Dharma-yanti INLP, et al. Serological Evidence of Ebola Virus Infection in Indonesian Orangutans. PLoS One [Internet]. 2012 Jul 18;7(7):e40740. Available from: https://doi.org/10.1371/journal.pone.0040740
- Soman Pillai V, Krishna G, Valiya Veettil M. Nipah Virus: Past Out-breaks and Future Containment. Viruses. 2020 Apr;12(4).
- Singh RK, Dhama K, Chakraborty S, Tiwari R, Natesan S, Khandia R, et al. Nipah virus: epidemiology, pathology, immunobiology and advances in diagnosis, vaccine designing and control strategies – a comprehensive review. Vet Q. 2019 Dec;39(1):26–55.
- Ang BSP, Lim TCC, Wang L. Nipah Virus Infection. J Clin Microbiol. 2018 Jun;56(6).
- Sendow I, Ratnawati A, Taylor T, Adjid RMA, Saepulloh M, Barr J, et al. Nipah Virus in the Fruit Bat Pteropus vampyrus in Sumatera, Indonesia. PLoS One [Internet]. 2013 Jul 22;8(7):e69544. Available from: https://doi.org/10.1371/journal.pone.0069544
- Sasaki M, Setiyono A, Handharyani E, Rahmadani I, Taha S, Adiani S, et al. Molecular detection of a novel paramyxovirus in fruit bats from Indonesia. Virol J [Internet]. 2012;9(1):240. Available from: https://doi.org/10.1186/1743-422X-9-240
- Sendow I, Field HE, Curran J, Darminto, Morrissy C, Meehan G, et al. Henipavirus in Pteropus vampyrus bats, Indonesia. Emerg Infect Dis [Internet]. 2 0 0 6 A p r ; 1 2 ( 4 ) : 7 1 1 – 2 . A v a i l a b l e f r o m : h t t p s : / /pubmed.ncbi.nlm.nih.gov/16715584
- Sendow I, Field HE, Adjid A, Ratnawati A, Breed AC, Darminto, et al. Screening for Nipah virus infection in West Kalimantan province, Indonesia. Zoonoses Public Health. 2010 Dec;57(7–8):499–503.
- Breed AC, Meers J, Sendow I, Bossart KN, Barr JA, Smith I, et al. The Distribution of Henipaviruses in Southeast Asia and Australasia: Is Wallace’s Line a Barrier to Nipah Virus? PLoS One [Internet]. 2013 Apr 24;8(4):e61316. Available from: https://doi.org/10.1371/journal.pone.0061316
- Wacharapluesadee S, Ghai S, Duengkae P, Manee-Orn P, Than-apongtharm W, Saraya AW, et al. Two decades of one health surveillance of Nipah virus in Thailand. One Heal Outlook [Internet]. 2021;3(1):12. Available from: https://doi.org/10.1186/s42522-021-00044-9
Most Commented